アポトーシスとは
個体が発生過程で臓器や組織を形作るときや古い細胞を新しい細胞に置き換えるとき、余分な細胞は死んでもらう必要があります。
たとえば胎児の手足にはカエルのようにみずかきがありますが、細胞が死ぬことで5本の指になります。
また、放射線や薬剤など外界からのストレスをうけたときや、細菌やウイルスに感染した時も、傷を受けた細胞は、増殖するより死んでもらうほうが個体が生きるのに都合がいいはずです。
このように、細胞は自爆装置を持っていて除かれるべきときに活性化していわゆる
自殺をします。
これが
アポトーシス(計画的細胞死)です。(後述遺伝子のミトコンドリアにも記述)
アポトーシスではDNAや細胞質は細切れになります。
細胞自身の分解酵素が働いて自分自身を分解して、マクロファージ(貪食細胞)に食べられやすくしているのです。
こうして死んだ細胞は内容物を外に漏れ出す前に貪食され、周りの正常な細胞に障害を与えることなく速やかに処理されます。
このことから、
アポトーシスは炎症を引き起こさない細胞死と一般的にいわれています。
それは、
マクロファージからIL10やTGF-βなどのサイトカインが放出され、周囲の炎症を抑制することにより、炎症反応を起こすことなくアポトーシス小体は組織から除去されるからです。
炎症反応は異物を破壊・除去し、損傷を修復することです
ウイルスや細菌などの異物が侵入すると、それを認識した
細胞から炎症誘導因子が放出され、血管が拡張し、発赤、熱感が現れたり、血管の透過性が亢進して腫脹が現れたりします。
また、異物と戦う
白血球が浸潤してきます。
これらは、
炎症性サイトカインを産生して他の白血球を活性化し、抗体を産生したり、異物の貪食(マクロファージ)を促進したり、感染細胞にアポトーシスを誘導したりします。
異物の除去が終わると、役目を終えた白血球は処理され、損傷を受けた組織は修復されます。
アポトーシスは炎症を誘導しない細胞死と前述しました。
たしかに、発生の段階などの細胞死で、その度に発赤や発熱などの炎症が起こっていたらたいへんなことです。
しかし、
特殊な状況ではあれ、アポトーシスが炎症を誘導する場合もあることもみつかってきました。
また、アポトーシスと炎症を分子レベルでみるとたいへん似ている処があるのです。
こうしてみると、アポトーシスと炎症は関係があるのではないかと考える事もできます。
アポトーシスの実行因子 カスパーゼ
アポトーシスのとき細胞質が細切れになることが重要です。
これには、細胞自身がもっている分解酵素が働いています。
この酵素はカスパーゼと呼ばれていて、細胞にアポトーシスを起こさせるシグナル伝達経路を構成する、一群のシステインプロテアーゼの総称です。
システインプロテアーゼは活性部位にシステイン残基をもつタンパク質分解酵素であり、カスパーゼは基質となるタンパク質のアスパラギン酸残基の後ろを切断します。
Caspaseという名はCysteine-ASPartic-acid-proteASEを略したものです。
カスパーゼは他のカスパーゼを切断し活性化するというカスケード(連鎖的増幅反応)の形で機能します。
また、
ある種のカスパーゼはサイトカイン(インターロイキン-1β)の活性化を通して免疫系の調節にも関与しています。
アポトーシスは正常な発生のほか、がんやアルツハイマー病などの疾病にも関係があることから、1990年代半ばに見出されて以来、治療のターゲットにもなりうるものとして注目されている。
カスパーゼは不活性な状態で合成されて、シグナルがきたときにだけ活性化します。
不活性な状態はプロドメインと呼ばれる領域がカスパーゼの活性化を抑制しているからで、シグナルがくるとこれらのドメインが切断されて活性化状態になります。
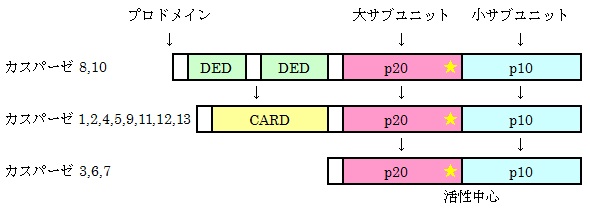
このような
プロドメインをもったものは上流カスパーゼといわれ、カスパーゼ1,2,4,5,8,9,10,11,12,13がこれに相当します。
プロドメインにはカスパーゼ調節因子が結合できる領域を持っていて
上流カスパーゼはこれら調節因子が結合することで自分自身を切断して活性化します。
たとえばカスパーゼ調節因子のひとつである
FADD(Fas-associated death domain)は
DED(death effector domain)とよばれる領域をもっていますが、FADDにも似たようなDED領域をもっていて、FADDのDED領域がカスパーゼ8のDED領域に結合してカスパーゼ8を活性化します。
このように似たような構造をもったものどうしが結合するのです。
また、
Apaf-1(apoptotic protease-activating factor 1)は
CARD(caspase recruitment domain)とよばれる領域をもった
カスパーゼ調節因子で、やはり似たようなCARD領域をもつ
カスパーゼ9に結合して活性化します。
これに対し下流カスパーゼはプロドメインが短いため調節因子に結合できず自分自身で活性化することができません。
これらのカスパーゼは上流カスパーゼによって切断されて活性化します。
カスパーゼ3,6,7,14がこれに相当します。
活性化した
下流カスパーゼはさまざまな基質を切断してアポトーシスを実行します。
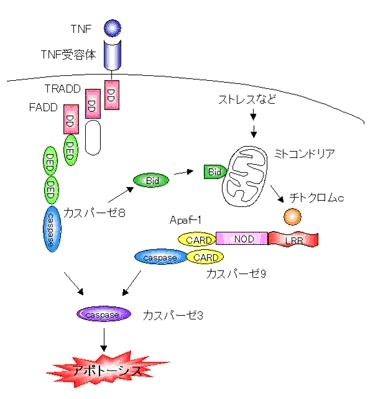
アポトーシス誘導メカニズム
紫外線や薬剤などのストレスをうけると、細胞内に存在するがん抑制遺伝子
p53が活性化して、細胞死を誘導するように働きかけます。
このときミトコンドリアからチトクロムcが流出します。
これがカスパーゼ調節因子であるApaf-1に結合して活性化しカスパーゼ9を活性化してアポトーシスを起こします。
また、
死受容体(death receptor・TNFやFas受容体)は細胞表面でシグナルをまっていて、免疫系などの細胞から提示される
TNFやFasリガンドなど(デスリガンド)に応答して細胞死を誘導します。
シグナルをうけた死受容体は
TRADD(TNF receptor-associated death domain)やFADD(Fas-associated death domain)といった
アダプター分子を介して
カスパーゼ8を活性化します。
カスパーゼ9やカスパーゼ8は上流カスパーゼであり自分自身を切断して活性化することを前に述べました。
Apaf-1やFADDはこれらのカスパーゼと結合し、カスパーゼ同士を近づけることによって切断し、活性化させる役割を果たしています。
アポトーシスを調節するBcl-2ファミリー
細胞内には、本当に死んでいいのか、生きるべきなのか判断する
アポトーシス制御因子が存在します。
Bcl-2ファミリータンパク質は、アポトーシスのシグナルのオン/オフを決定します。
Bcl-2ファミリーとはBH領域とよばれる領域をもった因子群のことをさし、これまでに30以上のタンパク質が見つかっています。
これらは、
アポトーシスを促進するほうに働くものと、
抑制するほうに働くものの大きく2つに分けられます。
両者の細胞内でのバランスによって細胞が生きるか死ぬかが決定されます。
Bcl2-like multidomain (アポトーシス抑制)
・Bcl2 antibodies、・Bcl-X (Bcl2L1) antibodies、・Bcl-W (Bcl2L2) antibodies
・Bfl1 (Bcl2A1) antibodies 、・MCL1 antibodies
Bax-like multidomain (アポトーシス促進)
・Bax antibodies、・Bak antibodies、・Bok antibodies
BH3-only (アポトーシス促進)
・Bid antibodies、・Bim antibodies、・Bad antibodies、・Bik antibodies
・Noxa antibodies、・Bmf antibodies、・Hrk antibodies、・BNIP3L antibodies
・PUMA antibodies
Bcl-2ファミリータンパク質はどのように調節されているのでしょうか
| 1)転写による量的調節 | インターロイキンなどの生存因子やSTAT、NF-kBなどの転写調節因子によってBcl-2ファミリータンパク質の発現が調節されています。 |
| 2)リン酸化や切断などの修飾による調節 | Bidはカスパーゼ-8によって切断されて活性化しますし、Badは通常リン酸化されていますが、脱リン酸化することで活性化します。 |
Bcl-2ファミリーは
ミトコンドリアからチトクロムcの流出を促進したり抑制したりすることでアポトーシスを制御します。
前述ようにチトクロムcが流出してApaf-1に結合するとカスパーゼ9を活性化してアポトーシスを起こします。
だとすると、FasやTNFを介したアポトーシスでは直接カスパーゼ8を活性化するのでBcl-2ファミリーの制御はうけないと思われますが、実際には、ミトコンドリアの経路を介してアポトーシスシグナルを増幅していると考えられています。
活性化したカスパーゼ8はアポトーシス促進型Bcl-2ファミリー因子であるBidを切断します。
これにより
Bidは活性になりミトコンドリアからチトクロムcを流出してアポトーシスを誘導します。
アポトーシスを抑制するClarp
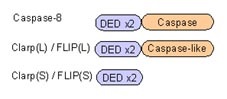
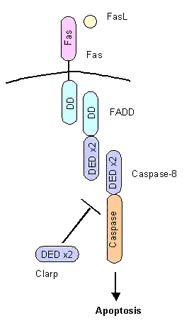
カスパーゼ8はそのDED領域を介してFADDのDEDに結合して活性化することは前に述べました。
このカスパーゼ8に似た分子が存在します。
Clarp(cFLIP)とよばれる因子です。
Clarpは
スプライシングによって短いものと長いものの二種類が存在。
Clarp(S型)はカスパーゼ8と似たようなDED領域を2つもちますが、カスパーゼ活性を示す領域をもっていないので、活性化することができません。
また、Clarp(L型)はカスパーゼ8と同じようにDED領域とカスパーゼ領域に似た領域をもっていますが、酵素活性をもっていません。
よって、ClarpはFADDやカスパーゼ8に結合しますが、アポトーシスを誘導することができません。
つまり、Clarpは「おとり」として働いており、カスパーゼ8を介したアポトーシスを抑制しているのです。
カスパーゼと炎症性サイトカイン
前述のとおり、カスパーゼはアポトーシス誘導にたいへん重要な因子です。
しかし、
カスパーゼはアポトーシスに関与するだけでなく、炎症反応にも関与しているのです。
炎症誘導に関与するものにはカスパーゼ1、5、11が含まれます。
カスパーゼ1と11は炎症誘導因子(炎症性サイトカイン)の産生に重要な働きをもちます。
ヒトのカスパーゼ5はマウスのカスパーゼ11に対応する遺伝子であると考えられています。
炎症性サイトカインは好中球の浸潤を誘導し炎症を起こします。
この炎症性サイトカインの代表的なものにIL-1βがあります。
通常IL-1βは不活性な状態で存在していています。
これはプロドメインが活性を抑制しているからで、この領域が切断されると活性型となって細胞外に放出されます。
別の炎症性サイトカインIL-18も同様の機構で活性化されます。
これらのプロドメインを切断するのがカスパーゼ1なのです。
カスパーゼ1の活性化にカスパーゼ11が必要であることもいわれています。
カスパーゼはアポトーシスを誘導するし、炎症性サイトカインの活性化も制御するのです。
アポトーシスと炎症のシグナル伝達経路
アポトーシスと炎症の関連性は他にもあります。
TNFによるアポトーシス誘導メカニズムについて前述しました。
TNFはアポトーシス誘導因子であり、炎症性サイトカインでもあります。
TNFがTNF受容体に結合すると受容体はTRADD、FADDとよばれるアダプター分子を介してカスパーゼ8を活性化し、アポトーシスを誘導します。
しかしその一方で、TRADDはRIPとよばれるアダプター分子を介してNF-kBの活性化も誘導します。
NF-kBはサイトカインの転写を活性化して炎症や免疫反応を起こします
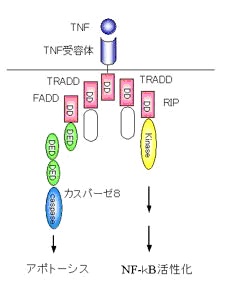
このことは、ひとつの
受容体からアポトーシス誘導のシグナルと炎症誘導のシグナルの2つのシグナルがでるということになります。




