ミトコンドリアの機能と働き
ミトコンドリア(mitochondrion pl. mitochondria)は真核生物の細胞小器官です。
二重の生体膜からなり、独自の
DNA(ミトコンドリアDNA=mtDNA)を持ち、分裂、増殖する。mtDNAはATP合成以外の生命現象にも関与し、酸素呼吸(好気呼吸)の場として知られている。
また、
細胞のアポトーシスにおいても重要な役割を担っているミトコンドリアDNAは、その遺伝子産物の一部が細胞表面にも局在し突然変異は自然免疫系が特異的に排除します。
ミトコンドリアは外膜と内膜という二枚の脂質膜に囲まれており、内膜に囲まれた内側をマトリックス、内膜と外膜に挟まれた空間を膜間腔と呼んでいる。内膜はマトリックスに向かって陥入しクリステ(稜)と呼ばれる特徴的な構造となっています。
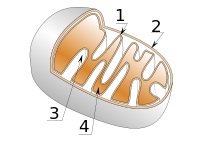
ミトコンドリアの構造
1.内膜 2.外膜 3.クリステ(平板状) 4.マトリックス
外膜:カルシウムシグナルの伝達や脂質の交換を行っています。
内膜:内膜はミトコンドリアの機能的アイデンティティを担っており、
酸化的リン酸化に関わる呼吸鎖複合体などの酵素群が規則的に配列しています。
呼吸鎖複合体は内膜をまたぐように
プロトン(水素イオン)勾配を形成し、それによって生じる膜電位が物質輸送や
ATP合成に関与しています。そのほかマトリックスへのタンパク質輸送装置やミトコンドリアの分裂・融合に関わるタンパク質群などが存在し、ミトコンドリアを構成する全タンパク質のおよそ2割(150以上)が含まれています。
マトリックス:内膜に囲まれた内側がマトリックスであり、
TCA回路(クレブス回路・
クエン酸回路)や
β酸化(脂肪酸を酸化して脂肪酸アシルを生成し、そこからアセチルCoAを取り出す代謝経路のこと)などミトコンドリアの
代謝機能に関わる酵素群が数多く存在しています。
ここには
ミトコンドリアDNA(mtDNA)が含まれており、
ミトコンドリア独自の遺伝情報が保持されています。その遺伝子発現を担うために、リボソーム、tRNA、転写因子や翻訳因子なども存在しています。ミトコンドリア全タンパク質の6~7割が存在しており、非常にタンパク質濃度の高い区画となっています。
ミトコンドリアの機能:ミトコンドリアの主要な機能は
電子伝達系による酸化的リン酸化によるATPの産生(ADPのリン酸化)です。
細胞のさまざまな活動に必要なエネルギーのほとんどは、直接、あるいは間接的にミトコンドリアからATPの形で供給されます。
しかしそれ以外にも多様な機能を持っており、
ステロイドやヘムの合成などを含む様々な
代謝、カルシウムや鉄の細胞内濃度の調節、細胞周期やアポトーシスの調節などにも大きく関わっているとされます。
ただし
すべてのミトコンドリアが上記の機能を担っている訳ではなく、あるものはある特定の細胞でのみ機能しています。
こうした様々な機能には多数の遺伝子が関わっており、それらに変異が自然免疫で排除されないとミトコンドリア病を引き起こすことになります。
ATP産生はミトコンドリアの主たる機能であって、これに関わる多くのタンパク質が内膜やマトリックスに存在しています。
細胞質には解糖系があり、グルコースを代謝することでピルビン酸とNADHを生じます。
もし酸素が十分に存在しない場合には解糖系の産物は嫌気呼吸により代謝されます。
しかしミトコンドリアで酸素を用いてこれらを酸化する好気呼吸を行うことで、嫌気呼吸と比べてはるかに効率よくATPを得ることができます。
嫌気性分解では1分子のグルコースから2分子のATPしか得られなかったのが、
ミトコンドリアによる好気性分解によって、1分子のグルコースから38分子のATPが合成できるようになった。
細胞質の解糖系で生成されたピルビン酸はピルビン酸共輸送体(ピルビン酸/H+)により細胞質からミトコンドリアへ輸送されます。
同じく細胞質で生成されたNADHはリンゴ酸-アスパラギン酸シャトルによりミトコンドリアへ実質的に輸送されます。
解糖系で生じたピルビン酸は内膜を能動輸送によって透過し、マトリックスで酸化され補酵素Aと結合し、二酸化炭素、アセチルCoA、NADHを生じます。
アセチルCoAはTCA回路へ入る基質です。
TCA回路の酵素群はほとんどがマトリックスに存在していますが、 コハク酸デヒドロゲナーゼ (ユビキノン)だけは例外で内膜の呼吸鎖複合体IIとなっています。
TCA回路はアセチルCoAを酸化して二酸化炭素を生じ、その過程で3分子のNADHと1分子のFADH2、1分子のGTPを生成します。
二酸化炭素はミトコンドリア外に排出されます。
アポトーシス(細胞死)とミトコンドリア
DNA損傷などのストレスは、アポトーシス誘導分子p53やアポトーシスを調節するBcl-2ファミリータンパク質を介して、ミトコンドリアの膜電位を変化させ、その結果、ミトコンドリアからチトクロムcが漏出し、アポトーシスへとつながります。
シトクロムcは、細胞質に存在する
Apaf-1や
カスパーゼ-9と結合して、アポトソーム(apoptosome)と呼ばれる集合体を形成し、これによって活性化されたカスパーゼ-9が、下流のエフェクターを活性化していきます。
がん(癌)とミトコンドリア
がんの発生とミトコンドリアDNA(mtDNA)の突然変異の関与は古くから指摘されてきました。
その理由は特定の発がん性化学物質がDNAよりもmtDNAに結合しやすい事、がん組織のmtDNAは正常組織よりも高い割合で突然変異が蓄積していた事が観察された事に依ります。
2008年筑波大学の林純一らが、
がんの転移能獲得という悪性化にミトコンドリアが関与している事を、マウス肺がん細胞の細胞質移植による細胞雑種の比較により、ミトコンドリアDNAの特殊な病原性突然変異によって
がん細胞の転移能獲得の原因になる事を発見し、ヒトのがん細胞株でもmtDNAの突然変異ががん細胞の転移能を誘導しえる事をあきらかにし、すくなくとも
mtDNAがATP合成以外の生命現象にも関与する事を明らかにしました。
また、林らによるとmtDNAの突然変異には
活性酸素種(ROS)の介在が重要であり、
ROSを除去すれば転移能の抑制が可能ではないかとしています。
がんの転移能の獲得メカニズムは複雑であり、様々な要因が考えられるのでこれはその要因の一つにすぎない。
2012年9月30日英国の科学誌「ネイチャー」(オンライン版) 良性腫瘍のがん化はミトコンドリアの機能低下が原因である事が神戸大学大学院医学研究科の井垣達吏・准教授らの研究で分かりました。
良性腫瘍は、複数の遺伝子が変異して生じた前がん細胞が過剰に増殖して作られますが、発生した場所から移動することはない。
この良性腫瘍が悪性化し、周囲の組織への浸潤や転移が起きるようになってがん(悪性腫瘍)になります。
がん化のメカニズム研究では、主にがん細胞での遺伝子変異が注目されてきたが、近年は、周辺細胞(ニッチ)との相互作用による影響も考えられるようになってきました。
井垣達吏・准教授らは、
良性腫瘍の中にある細胞でミトコンドリアの機能低下が起きると、その細胞から2種類のタンパク質(炎症性サイトカイン、細胞増殖因子)が産生、放出される。
これらによって、
周辺細胞内でがん抑制のために働くシグナル伝達経路(ヒポ〈Hippo〉経路)が阻害され、良性腫瘍が悪性化することが分かりました。